A wikipedia of Dr. D'Adamo's research
C O N T E N T SSee Also
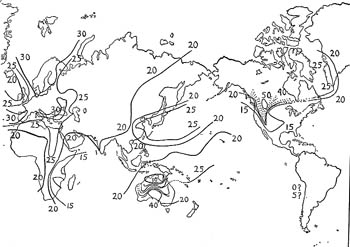
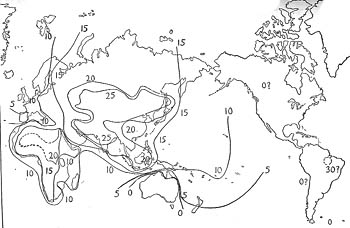
In the map (Figs. 1 and 2) are shown the results of blood grouping determinations on populations all over the world. Lines are drawn through points where the gene frequency is supposed to be the same. These lines are analogous with the isotherms and isobars which we find on weather maps, and can be referred to as isogenes. From them a picture of the geographical distribution of the three blood group genes may be obtained. Since we are not most interested in recent historical migrations, we have tried to avoid the confusing effect of recording data on recent migrants, and for America, the Pacific Islands, and Australia, we have plotted information only on the aborigines, omitting the results for the modern inhabitants, whose blood group frequencies, of course, are characteristic of their origins. We are not interested at present, for example, in the frequency of the four blood groups in the white inhabitants of Montana, but we are interested in the frequencies in the Indians who originally inhabited this state.
Fig.1: "Isogenes" for blood group gene A.
Fig.2: "Isogenes" for blood group gene B. The general situation is easily summarized. Looking at the isogenes for B, we notice that the highest incidence of B occurs in certain parts of Asia, and that in all directions as we go away from central Asia the incidence of group B declines. There is a subsidiary high center in Africa (and this seems to include the Nile Valley), although the frequency of q does not reach the highest levels we have seen in Asia. We find that the lowest figures in the Old World are reached in the Scandinavian countries, in the British Isles and the Spanish peninsula, where the percentage of the gene for B falls below per cent. In Australia the gene seems to have been absent until very recent times and there is every reason to believe that its introduction into a part of the northern coastal area is the result of fairly recent migration from the islands of the East Indies to the north and east. In the living aborigines of North America also we note a complete absence of the gene for B, except possibly in the Far North, and of course in Indians of part European ancestry group B is sometimes encountered. In South America the gene for B seems to be present only sporadically, if at all. One early report of a high percentage of group B in the Indians of Tierra del Fuego (1) has not been confirmed on retest (2,3). Another observation of B in South American Indians (4) was made with carefully checked reagents, but in view of the surprising nature of the results should nevertheless be redone. Certain Peruvian mummies, supposedly pre Columbian, seemed to possess the B factor (5), but more careful examination of the exact source of the specimens would be desirable before any sweeping conclusions are drawn from this observation. The map showing the incidence of the gene A presents a rather different situation. There is no large area of the world where A is absent, at least in the modern inhabitants. Among the North American Indians there are sharp differences from tribe to tribe. Some tribes are almost lacking in A; some tribes possess high percentages of A, and the Bloods and Blackfeet have percentages of A even higher than those possessed by any white population. It would hardly be possible or even desirable to discuss completely here the variations in blood group gene frequencies which exist in the various parts of the world which have been studied, but in addition to the general remarks already offered, we may mention a few results as typical. In Europe, for example, we find that in Germany the frequency of the B gene increases as we go from west to east, so that the Prussians of East Prussia have a higher frequency of group B (a typically Asiatic factor) than do the Jews of Berlin, in spite of whatever 'Semitic' (and thus Asiatic) ancestry the latter group may have (6). This increase in B frequency continues on into Poland and Russia. Conversely, the frequency of B falls off as we go west in the Spanish peninsula, and is subject to local variations such as a possibly complete absence of B in the Basques (7). In Italy the frequency of B increases as one goes from north to south (8, 9). In England a consistent gradient of the frequency of the gene A exists, with A increasing from north to south (10). The Japanese, before the Second World War, had already thoroughly determined the frequencies of the blood group genes throughout the home islands, and the accompanying map (Fig. 3 ) shows the data for the gene A.
Figure 3. The highest large concentration is in the area around the city Matsuye, on the island of Honshu, although there is another smaller concentration of A on the neighboring island of Kyushu. Going towards the north and east, the A concentration falls, although there is a secondary high center around Tokyo, and the lowest frequencies are reached in northeast Honshu, in the vicinity of the city of Morioka. The great regularity and fineness of the gradations in frequencies is only partly shown on the map reproduced here, which is based on a more complicated map in color published by Komatsu and Ito (11). If we think of mankind as having originated in Asia, it would look as if the gene A were possessed by the earliest wanderers from the original homeland, so that it is now widely dispersed, and on the whole common, although still a bit 'patchy' in its distribution. Gene B, on the other hand, is still most concentrated in the areas in which we think the human race may have had its origin. Early writers thought that this signified a late origin for B, but studies on anthropoids would suggest that B is no less ancient than A, and that it is largely accidental that the A gene spread so early and so widely, whereas the evidence clearly shows that B is in many places a recent importation. In all probability, both genes are older than the human species. The distribution of B in decreasing quantities around a center has led to its being called centripetal, and the wide-flung distribution of A has caused it to be called centrifugal (12). AttributionFrom: Genetics And The Races of Man, William C. Boyd. Little Brown and Company, Boston (1950) References1. Rahm, G., Invest. y Prog., 5, 160-162 (1931) 2. Lipschutz, A., G. Mostny, and L. Robin, AJPA, 4, 301-321 (1946) 3. Santiana, A., Los Fueguinos; sus grupos sangulneos, Quito, Imp. de la Univ., (1946) 4. Schiff, F., and H. Sasaki, Z. Imm., 77, 129-139 (1932) 5. Fischer, E., Die Rehobother Bastards. Verlag von Gustav Fischer, Jena, 1913. 6. Boyd, W. C., Quar. Rev. Biol., in press (1949) 7. Boyd, W. C., and L. G. Boyd, A. J. P. A., 23, 49-70 (1937) 8. Boyd, W. C., Quar. Rev. Biol., in press (1949) 9. Lattes, L., individuality of the Blood. Oxford Univ. Press, 1932 10. Schiff, F.A.J.P.A., 27, 255-262 (1940) 11. Komatso, Y., and R. Ito, Contrib. from Unit. Inst. Forensic Med., Kanazawa, 7 (1936) 12. Schiff, F., and W. C. Boyd, Blood Grouping Technic. Interscience Publishers, New York, 1942. |

COMPLETE BLOOD TYPE ENCYCLOPEDIA
The Complete Blood Type Encyclopedia is the essential desk reference for Dr. D'Adamo's work. This is the first book to draw on the thousands of medical studies proving the connection between blood type and disease. Click to learn more
Click the Play button to hear to Dr. Peter J. D'Adamo discuss .
|
The statements made on our websites have not been evaluated by the FDA (U.S. Food & Drug Administration).
Our products and services are not intended to diagnose, cure or prevent any disease. If a condition persists, please contact your physician.
Copyright © 2015-2023, Hoop-A-Joop, LLC, Inc. All Rights Reserved. Log In